

INTRODUCTION
Hand, Foot, and Mouth Disease (HFMD) is a common and typically self-limiting viral illness that predominantly affects infants and young children (1, 2). Globally recognized for its characteristic clinical presentation of fever, painful exanthems in the oral cavity, and a distinctive vesicular rash distributed across the palms, soles, and sometimes the gluteal region, HFMD poses a significant burden on pediatric healthcare systems and causes considerable distress to affected individuals and their families (3, 4). While the majority of cases resolve spontaneously within a week, infections with certain enteroviruses, notably Enterovirus 71 (EV71), are frequently associated with more severe and potentially life-threatening clinical outcomes, including a heightened risk of neurological complications such as aseptic meningitis, rhombencephalitis, brainstem encephalitis, and acute flaccid paralysis (1, 5, 6). These severe neurological sequelae can lead to long-term disabilities and, in some instances, result in fatal cardiopulmonary failure, underscoring the critical clinical importance of a comprehensive understanding of EV71 pathogenesis and the urgent need for the development of specific and effective antiviral therapies to mitigate its potentially devastating consequences in vulnerable pediatric populations (3, 7, 8).
The host-pathogen interface during viral infection is a critical disease progression and outcome determinant. Scavenger receptor class B2, also known as SCAB2 or CD36, emerges as a particularly versatile 53 kDa transmembrane glycoprotein, has traditionally been recognized for its roles in maintaining cellular homeostasis through the uptake of modified lipoproteins, such as oxidized low-density lipoprotein (oxLDL), and the clearance of cellular debris, including apoptotic cells (9). However, accumulating evidence has revealed that the functional repertoire of SRs extends significantly beyond these homeostatic functions to encompass the recognition and binding of a broad range of ligands, including a diverse array of microbial pathogens, such as bacteria, fungi, and viruses. By acting as pattern recognition receptors (PRRs), SRs play a crucial role in the early detection of invading microorganisms, contributing to the initiation of innate immune responses through the activation of downstream signaling pathways that lead to cytokine production and phagocytosis (9, 10). Understanding the specific SRs involved in the recognition of particular viruses is paramount for developing targeted antiviral strategies.
Given the established capacity of SCAB2 (CD36) to bind a diverse array of ligands, including certain bacterial and viral pathogens, we hypothesized that a soluble form of SCAB2, engineered for systemic circulation, could function as a decoy receptor for Enterovirus 71 (EV71) (10, 11, 12). By presenting the critical viral binding domain of SCAB2 in a soluble and readily accessible format, this decoy receptor could competitively bind to EV71 virions with high affinity, thereby effectively blocking their initial interaction with the native cellular receptor on susceptible target cells and consequently inhibiting the crucial early stages of viral infection, including attachment and entry. This novel therapeutic strategy offers a potentially advantageous approach to interfere with EV71 pathogenesis by specifically targeting the extracellular viral entry process, thus potentially circumventing issues associated with intracellular viral replication inhibitors, such as drug resistance and off-target effects on host cell machinery.
To rigorously investigate this hypothesis, the present study aimed to comprehensively evaluate the in vitro antiviral efficacy of a recombinant soluble form of SCAB2 fused to the human IgG1 Fc region (SCAB2:Fc) against a well-characterized strain of EV71 (13). Furthermore, we sought to elucidate the precise underlying mechanisms by which SCAB2:Fc exerts its observed antiviral effects. Our investigation focused on dissecting the impact of SCAB2:Fc on critical stages of the viral life cycle, including viral entry into host cells and subsequent intracellular viral replication. Additionally, we explored the potential modulation of key host cell signaling pathways, specifically those intricately involved in EV71 pathogenesis and the cellular response to viral infection, such as the mitogen-activated protein kinase (MAPK) signaling cascade. In this study, we intend to provide a robust scientific foundation for the potential development of SCAB2:Fc as a novel and targeted therapeutic intervention for effectively managing EV71-induced HFMD.
MATERIALS AND METHODS
Plasmid Construction and Protein Expression
The human Scavenger receptor class B (SCAB2) gene was cloned into the PCK plasmid using the restriction enzyme EcoRI. This plasmid was engineered to contain the human IgG1 Fc region downstream of the SCAB2 gene (SCAB2:Fc/PCK), enabling the production of a soluble protein (SCAB2:Fc) (13). The SCAB2:Fc/PCK plasmid DNA was then transfected into HEK293T cells, using a previously established protocol. Briefly, 1 µg of plasmid DNA was incubated with 7 µl of Polyethylenimine (PEI; Polysciences, Warrington, PA, USA) transfection reagent (5 mg/ml) for 15 minutes in a serum-free culture medium. This mixture was added to the cells and then incubated for 72 hours. The cell culture supernatant was collected and concentrated using a Centricon Amicon® Ultra Centrifugal Filter with a 30 kDa molecular weight cut-off cartridge (Merck Millipore Co., Burlington, MA, USA)). The resulting soluble SCAB2:Fc protein in the supernatant was quantified using a human IgG ELISA kit (Thermo Fisher Scientific, Waltham, MA, USA) (13).
Cells and Viruses
We obtained Enterovirus 71A (EV71A was provided by the Korea National Institute of Health) and amplified a live virus using HeLa cells. We grew HeLa cells for 16 hours and then infected them with EV71A (2). When more than 90% of the infected cells showed signs of cytopathic effect (CPE), we froze and thawed the cells three times at -80℃ to release the virus. To measure the amount of virus in our stock, we used a TCID50 assay. In this assay, we made a series of dilutions of the virus and added each dilution to several wells of HeLa cells. After a certain time, we looked for CPE in each well. The TCID50 is the virus concentration that causes CPE in 50% of the wells. We calculated this using a standard statistical method. We cultured the HeLa cells in a specific growth medium (Dulbecco’s Modified Eagle Medium, or DMEM) that contained 5% fetal bovine serum (FBS) and 1% penicillin-streptomycin (from Welgene, Inc. in Gyeongsan, Korea). We incubated these cells at 37℃ in a humidified environment with 5% CO2.
Western blot analysis
Protein was extracted using 1X PBS lysis buffer (1X PBS, 0.5 mM EDTA, 0.1% Triton X-100, and protease inhibitor cocktail). The cell lysate was centrifuged at 4℃, 13000 rpm, for 5 minutes. 1 Aliquots of the total cell extract supernatant were loaded onto 10% SDS-polyacrylamide gel electrophoresis (SDS-PAGE) gels. After electrophoresis, the cells were transferred to polyvinylidene difluoride (PVDF) membranes. The membranes were blocked in 5% non-fat dry milk solution in Tris-buffered Saline containing 0.1% Tween 20 (TBST) and probed with 2 anti-SB2, EV71-VP1, eukaryotic translation initiation factor 4 Gamma 1 (eIF4G1) (1:1000, rabbit polyclonal antibody), glyceraldehyde-3-phosphate dehydrogenase (GAPDH) (1:2500, mouse monoclonal antibody), Phospho-Akt (Ser473) (1:2500, mouse monoclonal antibody), phospho-p38 mitogen-activated protein kinase (MAPK) (Thr180/Tyr182), Phospho-GSK-3β (Ser9), phospho-nuclear factor kappa-light- chain-enhancer of activated B cells (NF-κB), total Akt, total p38 MAPK, total GSK-3β, and total NF-κB (1:1000, rabbit polyclonal antibody) (Cell signaling, MA, USA) for overnight at 4℃ and then detected by the Chemi-doc system (Bio-Rad, CA, USA) after ECL plus solution (Intron Biotech, Sengnam, Korea) treatment.
Immunofluorescent Stain
We confirmed the proliferation of EV71A by immunofluorescent staining of the capsid protein VP1. HeLa cells were infected with EV71A after a 30-minute pre-incubation with serially diluted sSCAB2 protein. Sixteen hours post-infection, the HeLa cells were fixed with cold methanol for 15 minutes. Subsequently, the cells were blocked and permeabilized with a blocking solution of 2% Bovine Serum Albumin (BSA) and 0.2% Triton X-100 in PBS. The cells were then incubated with a rabbit EV71 VP1 primary antibody (Merck Millipore, Germany). The target proteins were visualized using secondary antibodies conjugated with fluorophores (Alexa Fluor 488, 1:250; Merck Millipore, MA, USA). Fluorescence images were captured and processed using a fluorescent microscope (Olympus Co., Japan). The fluorescence intensity in the images was quantified using NIH-ImageJ software (14).
Reverse Transcription qPCR
Total RNA was isolated from virus-infected samples using the TRIzol reagent (ThermoFisher Scientific, Cambridge, MA, USA). Complementary DNA (cDNA) was synthesized using 1ug RNA through a reverse transcription reaction using an oligo-dT primer for RNA quantification. Real-time PCR quantitative RNA or DNA analyses were performed in an ABI Sequence Detection System using the SYBR green fluorescence quantification system (Applied Biosystems, Waltham, MA, USA). EV71A positive- and negative- RNA amplification was measured by real-time PCR, respectively. PCR with the synthesized cDNA was performed with EV71-VP1 and GAPDH primers. EV71 sense primer (5’-GTTCTTAACTCACATAGCA-3’), and EV71 antisense primer (5’-TTGACAAAAACT- GAGGGGTT-3’); GAPDH sense primer (5’-ATCAACGACCCCTTCATTGAC-3’), and GAPDH antisense primer (5’-CCAGTAGACTCCACGACATACTCAGC-3’).
Statistical analysis
All data was analyzed using Prism 9 software (GraphPad Software, San Diego, CA, USA), and the results are presented as the means ± standard deviation (SD). To determine the statistical significance between two groups, we used a Student’s t-test. For comparisons involving more than two groups, we performed a one-way ANOVA. Statistical significance was defined as *p < 0.05, **p < 0.01, and ***p < 0.001.
RESULTS
Generation and characterization of SCAB2:Fc protein
Soluble SCAB2:Fc protein was generated from HEK293T cells by SCAB2:Fc/PCK plasmid transfection. After 72 hours of transfection, the protein production was confirmed by Western blot analysis in the cultured cell lysate. Fig. 1A shows the design of SCAB2:Fc/PCK for protein production. Present data confirming the successful transfection of HEK293T cells and the secretion of functional SCAB2:Fc proteins by western blot analysis. The membrane was probed by human IgG-HRP and SR-B2 antibodies (Fig. 1B). The concentration of SCAB2:Fc protein in the cell supernatant was measured by human IgG ELISA-based quantification.
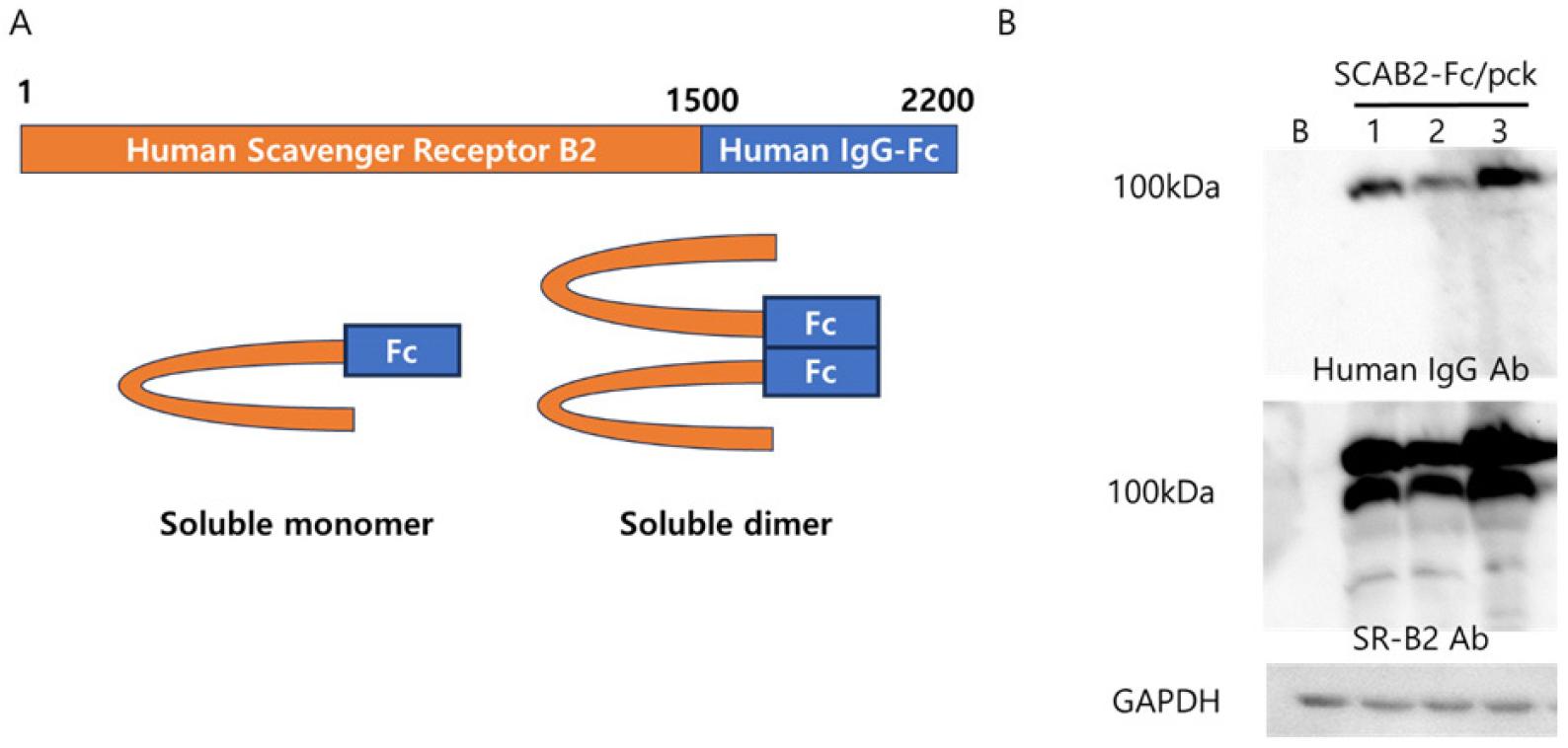
Fig. 1
Generation of soluble SCAB2:Fc plasmid and confirmation of protein production. (A) The diagram of soluble SCAB2: Fc/PCK plasmid. Human scavenger receptor type B2 cDNA was inserted into the human IgG-Fc/PCK plasmid. (B) soluble SCAB2:Fc protein production was observed from SCAB2:Fc/PCK plasmid-transfected HEK293T cells. After 72 hours, the total protein was extracted and then applied for western blot analysis, then proved human IgG-HRP antibody, scavenger receptor B2 (SR-B2) antibody, and GAPDH antibody.
SCAB2:Fc blocks EV71A infection
The antiviral effect of newly generated SCAB2:Fc protein was observed by immunofluorescent stain assay (IFA) after EV71A co-incubation for 18 hours. Increasing concentrations of SCAB2:Fc lead to a significant reduction in EV71A entry into susceptible HeLa cells. The virus particle was detected by EV71A capsid protein VP1 antibody (Fig. 2A). The present quantitative data support the dose-dependent blocking effect of SCAB2:Fc for EV71A (Fig. 2B). These results suggest the strong antiviral effects of SCAB2:Fc protein, which successfully inhibits EV71A infection.
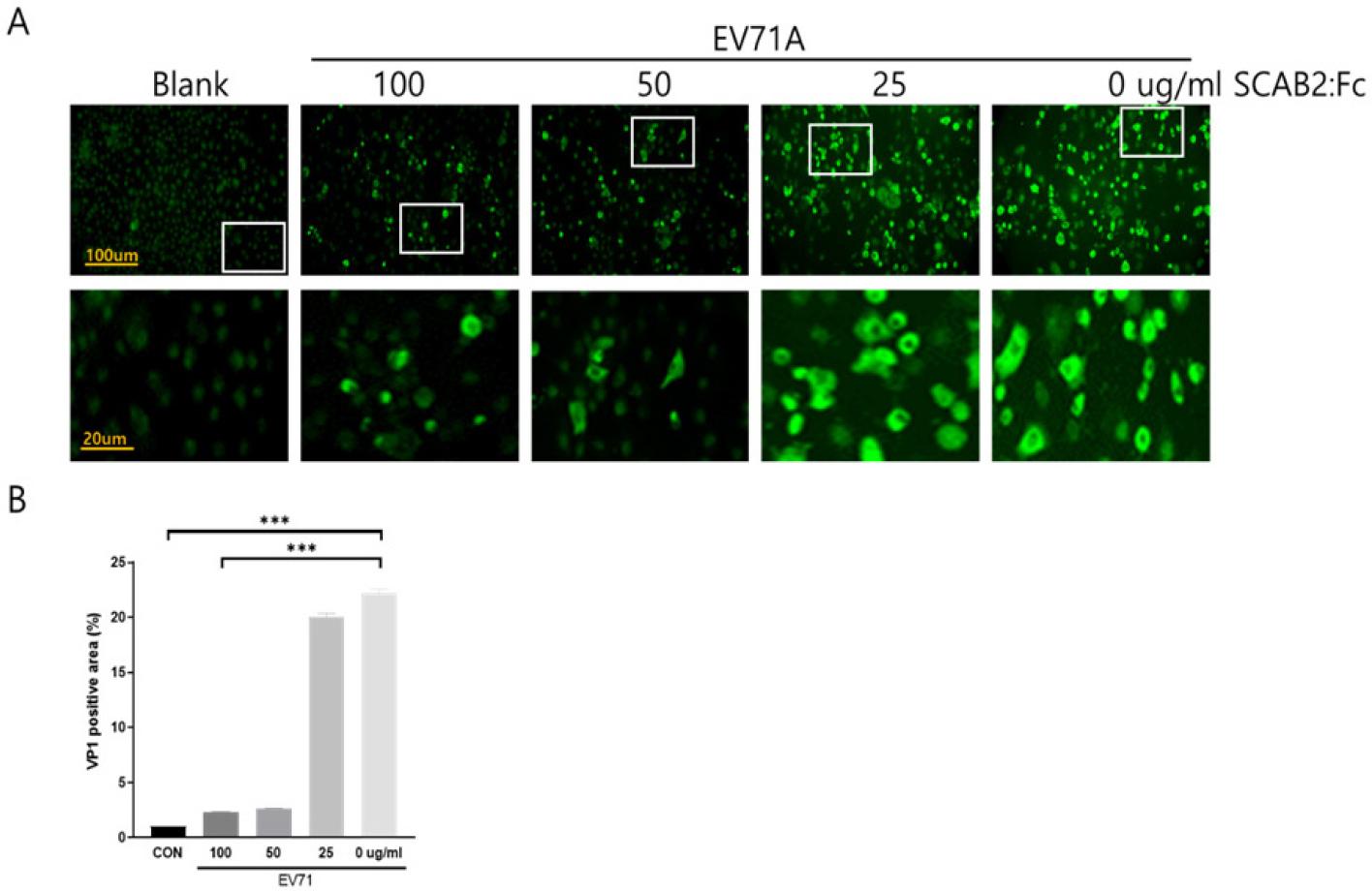
Fig. 2
SCAB2:Fc Blocks EV71A infection. (A) The virus-blocking effects of soluble SCAB2:Fc were observed in EV71A cell infection by immunofluorescence stains. EV71A capsid protein VP1 was stained green fluorescent. (B) The inside cell virus capsid VP1 protein was stained and quantified by NIH-imageJ software. Data are expressed as the mean ± SD, *** P<0.001 by a two-tailed Student’s t-test.
SCAB2:Fc inhibits EV71A replication
The inhibition effects of SCAB2:Fc were observed by virus capsid protein VP1 production and virus gene amplification change. Western blot analysis showed a significant decrease in the expression of the VP1 capsid protein in cells treated with SCAB2:Fc compared to untreated cells (blank). VP1 protein expression was significantly decreased in a dose-dependent manner. 25ug/ml of SCAB2:Fc significantly inhibited the virus replication in infected HeLa cells (Fig. 3A). The VP1 protein expression was quantified by NIH-ImageJ software (Fig. 3B). Viral gene expression is an important indicator for virus replication and progeny virus production. The results from reverse transcription PCR (RT-PCR) demonstrated a significant reduction of EV71A positive-strand RNA gene in SCAB2:Fc-treated cells (Fig. 3C) compared to untreated cells. Provide quantitative data and statistical significance for VP1 protein and viral positive-strand RNA reduction. These results strongly support that the soluble SCAB2:Fc protein successfully inhibits EV71A infection and replication.
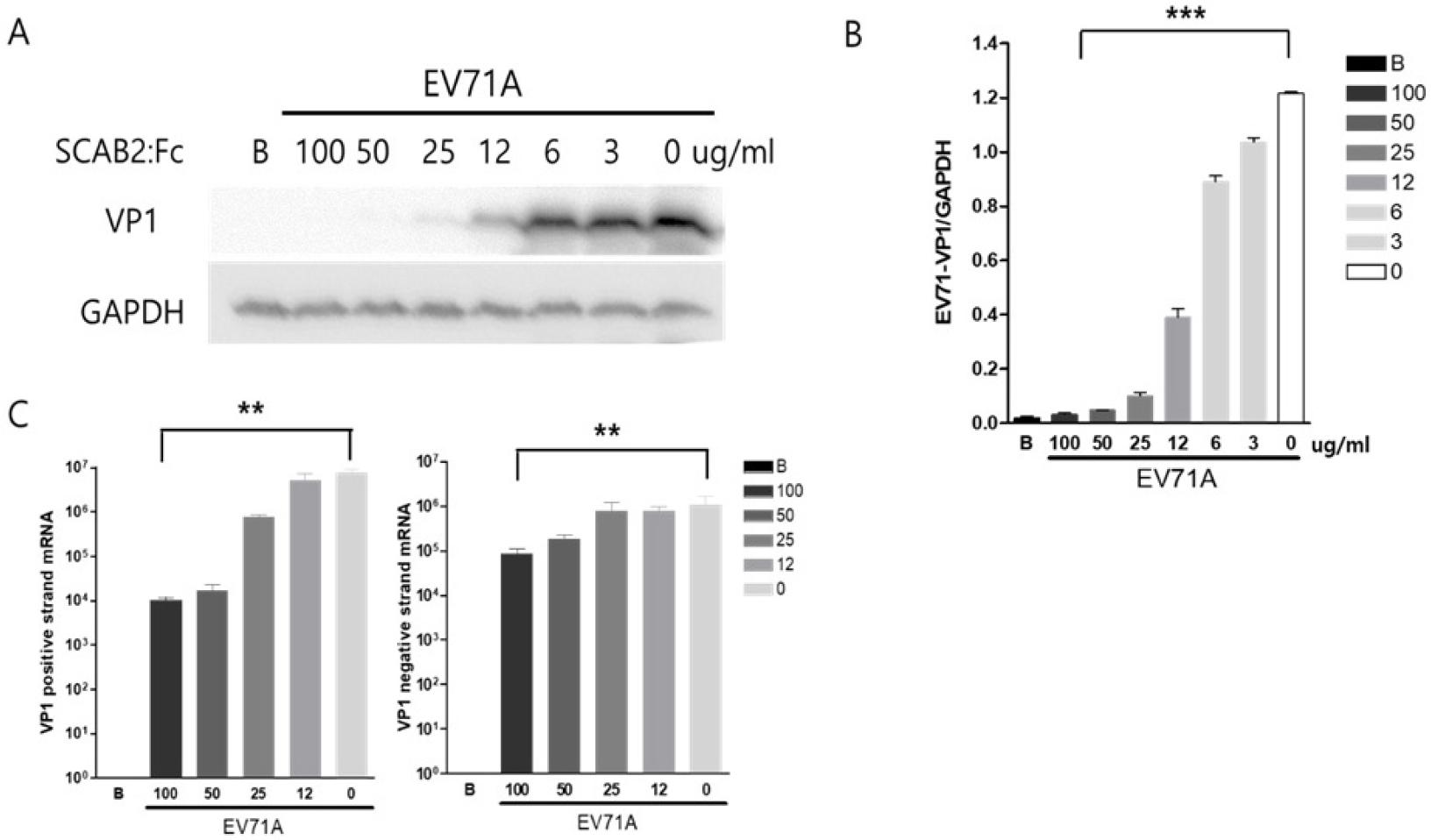
Fig. 3
SCAB2:Fc decreases EV71A replication. (A) After 18 hours, the protein was extracted from the cells with or without soluble SCAB2:Fc in EV71A infection and then applied for western blot analysis probed by EV71A-VP1 antibody. (B) Western blot result was quantified using NIH-imageJ software. (C) Total RNA was extracted from treated HeLa cells and then applied for real-time PCR quantified EV71A VP1 positive and negative strand mRNA. Data are expressed as the mean ± SD, ** P<0.01, *** P<0.001 by a two-tailed Student’s t-test.
SCAB2:Fc decreases cell apoptosis in EV71A infection
Virus infection is a well-established inducer of host gene expression inhibition and apoptosis signaling in infected cells. This study investigated the protective efficacy of SCAB2:Fc by examining its impact on key apoptotic regulators, specifically caspase-3 and p62. We observed a notable reduction in the protein levels of the translation initiation factor eIF4G1 in SCAB2:Fc-treated cells compared to untreated, virus-infected controls. We showed that SCAB2:Fc treatment led to an upregulation of the autophagy receptor and apoptosis inhibitor p62, accompanied by a reduction of the cleaved caspase-3, a critical executioner of apoptosis (Fig. 4A and B). These findings collectively suggest that SCAB2:Fc exerts a protective effect against virus-induced apoptosis. Furthermore, pre-treatment with SCAB2:Fc demonstrated a dual inhibitory action, effectively blocking viral entry and suppressing apoptosis triggered by viral infection. These results highlight the potential of SCAB2:Fc as a therapeutic intervention against EV71A pathogenesis involving apoptosis.

Fig. 4
Soluble SCAB2:Fc treatment reduced EV71A-infected cell apoptosis. (A) Protein was extracted from the cells with or without soluble SCAB2:Fc in EV71A infection and then applied for western blot analysis probed by eIF4G1, p62, and cleaved caspase3 antibodies. (B) Western blot result was quantified using NIH-imageJ software. Data are expressed as the mean ± SD, ** P<0.01, *** P<0.001.
SCAB2:Fc treatment reduces phosphorylation levels of p38 and ERK1/2 in EV71A-infected cells
We want to define the main cell signaling of EV71A antiviral effects. Western blot analysis revealed a significant decrease in the phosphorylation levels of p38 and ERK1/2 in EV71A-infected cells treated with SCAB2:Fc compared to untreated infected cells (Fig. 5A). Quantitative analysis demonstrated a statistically significant reduction (p < 0.05) in the ratio of phosphorylated to total p38 and ERK1/2 protein levels in the SCAB2:Fc-treated group, suggesting that SCAB2:Fc inhibits the activation of the p38 and ERK1/2 signaling pathways induced by EV71A infection. The expression levels of total p38 and ERK1/2 proteins were used as loading controls (Fig. 5B).
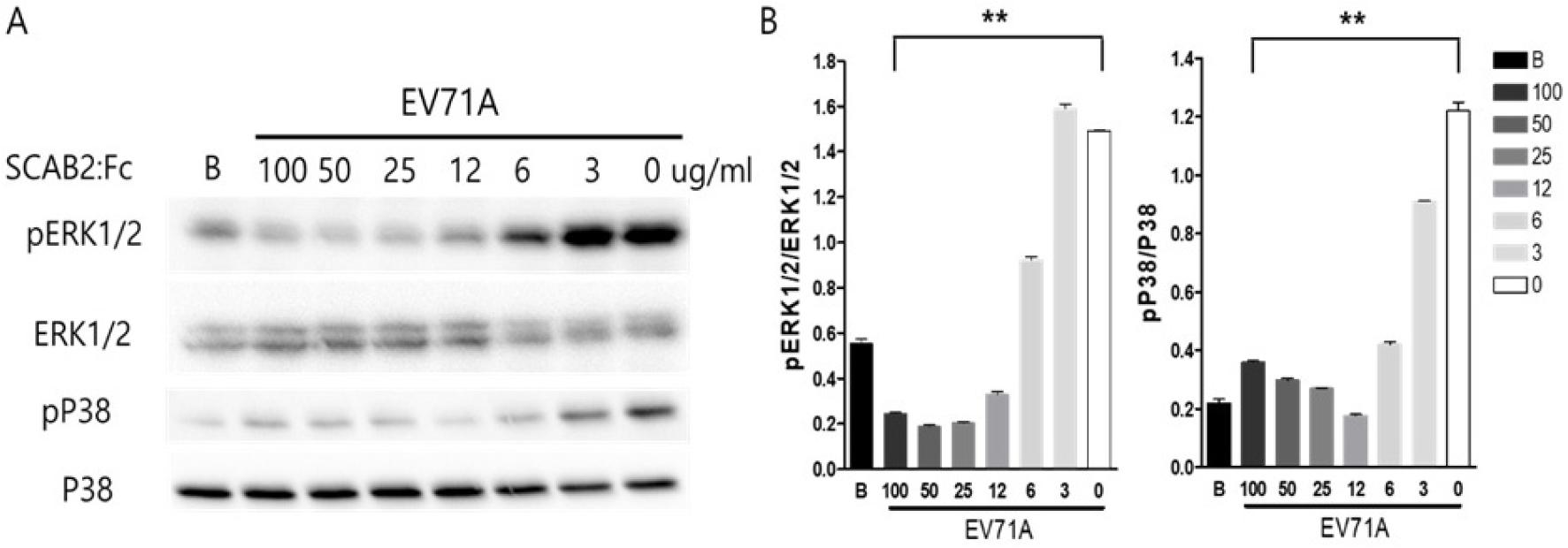
Fig. 5
Soluble SCAB2:Fc treatment decreases EV71A replication through the inhibition of MAPK signaling cascade. (A) Protein was extracted from the HeLa cells after soluble SCAB2:Fc treatment with EV71A infection and then applied for western blot analysis probed by phospho-ERK1/2, total ERK1/2, phosphor-p38, and total p38 antibodies. (B) Western blot result was quantified using NIH-imagyJ software. Data are expressed as the mean ± SD, ** P<0.01, *** P<0.001.
DISCUSSION
Our study demonstrated that a soluble viral receptor protein called SCAB2:Fc, which we produced by introducing specific DNA into cells, effectively neutralized enterovirus 71A (EV71A) in the cell culture. Furthermore, this manufactured SCAB2:Fc protein showed the potential to treat EV71A infection. It reduced EV71A infection and the virus’s ability to multiply by blocking cell signaling pathways (p38 and ERK1/2 phosphorylation) that typically lead to cell death (apoptosis) and virus replication. Essentially, this protein helped infected cells survive and stopped the virus from making more copies of itself.
Hand, Foot, and Mouth Disease (HFMD) is a common viral illness, usually resolving on its own, that primarily affects babies and young children (1). It’s recognized worldwide by its typical symptoms: fever, painful sores in the mouth, and a distinctive rash with small blisters on the palms of the hands, soles of the feet, and sometimes the buttocks. HFMD places a significant strain on children’s healthcare and causes considerable distress for those affected and their families (2, 3). While most cases get better on their own within a week, infections with certain enteroviruses, particularly EV71A, are often linked to more severe and potentially life-threatening complications. These include a higher risk of neurological issues like aseptic meningitis, rhombencephalitis, brainstem encephalitis, and acute flaccid paralysis (1, 3, 7). Recently, there have been no fully developed therapeutic drugs for this.
Previously, we reported on several potential antiviral drugs derived from natural sources that could be developed as new treatments for EV71A. In a separate study, we examined the antiviral activity of an extract from Streptomyces sp. zx10-19 (KH29) against EV71A (15). Treating cells infected with EV71A with KH29 extract (at a concentration of 100 μg/ml) significantly reduced the production of the EV71A capsid protein VP1 and the breakdown of the translation initiation factor eIF4G1 within these cells. Additionally, the KH29 extract significantly inhibited the production of the positive and negative strands of the EV71A viruses RNA. Moreover, we observed salvianolic Acid B (SalB) extract (2) treatment, significantly increased the activation of the PKB/AKT signaling pathway in EV71A-infected cells. This suggests that SalB extract can help cells survive by activating the AKT signaling pathway and effectively prevent EV71A from multiplying. While both SalB and KH29 extract activate the Akt signaling pathway (16, 17, 18), KH29 extract appeared to have a broader antiviral effect by interfering with the early stages of viral RNA production. However, these natural compounds did not completely stop EV71A from entering cells. In contrast, the soluble SCAB2:Fc protein initially and dramatically blocked EV71A from entering cells and significantly reduced virus replication.
Furthermore, enteroviruses multiply by activating cell signaling molecules called ERK1/2 and p38 (19, 20, 21, 22). Our findings also showed that the activity of MAPK (mitogen-activated protein kinases) and p38 was significantly reduced after we administered the soluble SCAB2:Fc. As a result, the cells remained alive because the protein prevented the virus from triggering cell death. This is a crucial finding, suggesting that SCAB2:Fc could be used to develop therapeutic drugs for EV71A. However, we haven’t conducted animal studies yet because there isn’t a well-established animal model for Enterovirus 71A. This lack of a suitable model limits our ability to perform in vivo experiments. Therefore, future research should focus on creating an effective animal model and then using it to test how well potential treatments work.
This treatment strategy offers a potentially beneficial way to fight EV71A by specifically targeting how the virus enters cells from the outside. This approach might avoid problems often seen with drugs that work inside cells, such as the virus developing resistance and the drug affecting normal cell functions. Moreover, we understand exactly how SCAB2:Fc produces its antiviral effects. Our research suggested that SCAB2:Fc affects critical steps in the virus’s lifecycle, including how it gets into host cells and replicates. These findings demonstrated that further investigation into the effectiveness and potential use of soluble SCAB2:Fc in living organisms could lead to a new treatment for HFMD caused by EV71A.
