

INTRODUCTION
Vibrio vulnificus(패혈증비브리오균)은 호염성의 그람음성막대균으로, 오염된 해산물을 섭취하거나, 피부상처가 오염된 해수에 노출되어 감염된다 (1, 2). 감염 사례는 계절적 분포가 뚜렷하여 대부분 초여름부터 늦가을 사이에 발생한다 (3). 사람 간 전파는 일어나지 않으며, 만성 간질환자, 면역저하환자 등이 고위험군이며, 치사율은 약 50%로 높은 사망률을 보인다 (4, 5). 우리나라에서는 1979년 전라남도에서 발생한 원인을 알 수 없는 피부괴질이 1983년 패혈증비브리오균 감염에 의한 것으로 밝혀지면서 처음으로 보고되었다 (6). 우리나라는 2000년 비브리오패혈증을 3급 법정감염병으로 지정하였으며, 이후 2023년까지 전국에서 1,285명의 비브리오패혈증 환자가 발생하였으며, 그중 전라남도가 227명(17.7%)으로 지역별 발생건수가 가장 높다 (7).
그동안 패혈증비브리오균의 병원성을 예측하기 위한 생화학적, 유전학적 다양한 시도들이 수행되었다 (8, 9, 10). 분자유전학적 접근 가운데 분리 출처에 따라 패혈증비브리오균의 16S rRNA(rrn) 염기서열에 차이가 존재한다는 것이 알려졌다 (11). 또한 random-amplified polymorphic DNA polymerase chain reaction (RAPD-PCR)을 통해 유전학적 표지자로 독성상관유전자(virulence-correlated gene, vcg)가 확인되었다 (12, 13). 대부분의 임상분리균주에서는 rrnB와 vcgC가, 환경분리균주에서는 rrnA와 vcgE가 확인되므로 임상형 유전형(rrnB/vcgC)을 가진 균주가 독성이 더 강할 것으로 추정되었지만 (14), 생쥐모델 실험을 통한 독성연구에서는 패혈증비브리오균의 유전형과 독성의 상관성이 명확하지 않은 결과를 나타내었다 (15, 16). 그럼에도 독성에 대한 신뢰할 수 있는 표지자가 아직까지는 불분명하므로 유전형에 의한 분류법이 지속적으로 이용되고 있다.
국내 패혈증비브리오균 연구는 임상적, 역학적 분석 (3, 17), 독소 유전자의 분포 및 항생제 내성에 관한 연구 (18, 19), 그리고 환경인자와의 연관성 확인 연구가 대부분을 차지하고 있다 (20, 21). 분자 역학적 방법 중 하나인 pulsed-field gel electrophoresis (PFGE)로 패혈증비브리오균을 분석하면 높은 유전적 다양성이 확인되고, 환경분리균주의 지역적인 상동성 및 유사성은 낮다 (22, 23). 또 다른 분자생물학적 역학조사 방법인 multi-locus sequence typing (MLST)는 house-keeping 유전자의 염기서열을 이용하여 균주 간의 차이를 분석하는 방법이다 (24). MLST는 무작위적인 유전자 변이를 젤 전기영동을 통해 구분하는 PFGE와 비교하여 명확한 염기서열 정보로 각 균주의 유전자 정보를 쉽게 비교할 수 있으며, 밀접한 유연관계의 균주들을 구별할 수 있어 패혈증비브리오균의 계통도 분석에 이용된다.
본 연구는 전라남도 보건환경연구원에서 진단한 임상분리균주 35주와 서해안 지역에서 분리한 환경분리균주 42주 등 총 77주의 패혈증비브리오균을 대상으로 유전형 확인과 MLST 분석을 수행하여 균주의 계통학적 위치와 그 특성을 분석하였다. 이를 통해 임상분리균주와 환경분리균주들 간의 계통학적 관계를 바탕으로 향후 우리나라에서 분리되는 패혈증비브리오균의 분자 역학적 연구에 기초자료를 제공하고자 한다.
MATERIALS AND METHODS
균주
연구에 사용된 패혈증비브리오균은 총 77주로 임상분리균주는 9년간(2014-2022년) 전라남도 지역 의료기관에서 비브리오패혈증으로 의뢰하여 전라남도 보건환경연구원에서 진단한 35주를 사용하였다 (Table 1). 환경분리균주는 2022년 연중, 월 2회 전라남도 서해안 지역(영광, 함평)의 해수 및 갯벌에서 분리한 37주와 2020년과 2021년에 저온(3.7~8.0℃)의 해수에서 분리한 5주를 포함하여 총 42주를 사용하였다 (Table 2).
Table 1.
Characteristics of V. vulnificus isolates from clinical samples
Table 2.
Characteristics of V. vulnificus isolates from environmental samples
| Isolate No. | Collection year | Collection month | Sample source | rrn typing | vcg typing | Sampling sitea |
| JNIHE_E1 | 2022 | Feb | Seawater | B | C | YG |
| JNIHE_E2 | 2022 | Mar | Seawater | B | C | YG |
| JNIHE_E3 | 2022 | Mar | Sea mud | B | C | YG |
| JNIHE_E4 | 2022 | Apr | Seawater | B | C | YG |
| JNIHE_E5 | 2022 | May | Seawater | B | C | YG |
| JNIHE_E6 | 2022 | May | Seawater | B | C | YG |
| JNIHE_E7 | 2022 | Jun | Seawater | B | C | YG |
| JNIHE_E8 | 2022 | Jun | Seawater | B | C | HP |
| JNIHE_E9 | 2022 | Jun | Seawater | B | C | YG |
| JNIHE_E10 | 2022 | Jun | Sea mud | B | C | YG |
| JNIHE_E11 | 2022 | Jun | Seawater | A | E | HP |
| JNIHE_E12 | 2022 | Jun | Sea mud | B | C | HP |
| JNIHE_E13 | 2022 | Jul | Seawater | B | C | YG |
| JNIHE_E14 | 2022 | Jul | Sea mud | B | C | YG |
| JNIHE_E15 | 2022 | Jul | Sea mud | - | C | HP |
| JNIHE_E16 | 2022 | Jul | Sea mud | B | E | YG |
| JNIHE_E17 | 2022 | Jul | Sea mud | B | C | HP |
| JNIHE_E18 | 2022 | Aug | Seawater | B | C | YG |
| JNIHE_E19 | 2022 | Aug | Seawater | B | C | HP |
| JNIHE_E20 | 2022 | Aug | Sea mud | B | C | HP |
| JNIHE_E21 | 2022 | Aug | Seawater | B | C | YG |
| JNIHE_E22 | 2022 | Aug | Seawater | B | C | HP |
| JNIHE_E23 | 2022 | Aug | Sea mud | B | C | HP |
| JNIHE_E24 | 2022 | Sep | Seawater | B | C | YG |
| JNIHE_E25 | 2022 | Sep | Sea mud | B | C | YG |
| JNIHE_E26 | 2022 | Sep | Seawater | A | E | HP |
| JNIHE_E27 | 2022 | Sep | Sea mud | B | C | YG |
| JNIHE_E28 | 2022 | Sep | Seawater | A | E | HP |
| JNIHE_E29 | 2022 | Oct | Seawater | B | C | YG |
| JNIHE_E30 | 2022 | Oct | Sea mud | B | C | HP |
| JNIHE_E31 | 2022 | Oct | Seawater | A | E | YG |
| JNIHE_E32 | 2022 | Oct | Sea mud | - | C | YG |
| JNIHE_E33 | 2022 | Oct | Seawater | B | C | HP |
| JNIHE_E34 | 2022 | Nov | Seawater | B | C | YG |
| JNIHE_E35 | 2022 | Nov | Sea mud | B | C | YG |
| JNIHE_E36 | 2022 | Nov | Seawater | B | C | YG |
| JNIHE_E37 | 2022 | Dec | Seawater | B | C | YG |
| JNIHE_E38 | 2020 | Jan | Seawater | B | C | YG |
| JNIHE_E39 | 2020 | Jan | Seawater | B | C | YG |
| JNIHE_E40 | 2020 | Feb | Seawater | B | C | YG |
| JNIHE_E41 | 2020 | Feb | Seawater | B | C | YG |
| JNIHE_E42 | 2021 | Jan | Seawater | - | C | YG |
검체 관련 정보 수집
본 연구에 활용된 임상검체 관련 정보는 검체 시험의뢰서의 인적 특성(성별, 나이)을 활용하였다. 같은 기간(2014-2022년) 전라남도에서 발생한 비브리오패혈증 69건의 인적 특성은 전라남도의 성별/연령별 감염병통계를 이용하였다 (7). 환경검체 관련 정보는 검체 채취 시 측정한 해수 및 갯벌의 온도를 이용하였다.
균 분리를 위한 배양 및 균종 동정
임상검체는 tryptic soy broth (TSB, Oxoid, UK)에 접종하여 37℃에서 16시간 증균 배양하였다. 환경검체는 1 m 이내의 표층수 250 mL 해수를 여과한 0.45 µm여과막(Advantec, Japan) 또는 약 2.5 g의 갯벌을 alkaline 펩톤수(pH 8.6)에 접종하여 37℃에서 하룻밤 정치 배양하였다. 패혈증비브리오균의 분리를 위해서는 배양액을 thiosulfate citrate bile-salt sucrose (TCBS, Oxoid, UK)배지에 도말하여 녹색집락을 확인한 후, CHROMagar-Vibrio (CHROMagar, France)배지에서 청록색을 나타내는 집락을 선택하여 1% NaCl이 첨가된 tryptic soy agar (TSA, Oxoid, UK)배지에 순수 분리하였다. 최종적으로 VITEK 장비(Biomerieux, France)를 이용하여 생화학적 동정을 실시하여 신뢰도 90% 이상되는 균주를 사용하였다.
유전형 확인을 위한 PCR
순수 배양된 단일집락을 멸균된 증류수 200㎕에 현탁한 후, NX-48 Bacterial DNA Kit (Genolution, Korea)와 자동화 핵산추출장비 NX-48 (Genolution, Korea)를 이용하여 DNA를 추출하였다. 유전형 확인을 위해 16S rRNA유전자와 독성상관유전자(vcg)를 증폭하였으며, 이전 연구에서 사용된 primer와 반응조건을 사용하였다 (12, 25). PCR 반응은 AccuPower PCR PreMix (Bioneer, Korea) 튜브에 각 primer 1㎕와 추출된 DNA 2㎕를 첨가한 뒤 증류수로 최종 부피를 20㎕로 만들어 SimpliAmp Thermal Cycler (Thermo Fisher Scientific, USA)기기를 이용하여 수행하였다. 유전자 증폭을 확인하기 위해 PCR 산물 4㎕를 1.5% agarose gel에 40분간 전기영동 하였다. 두 유전자를 통해 패혈증비브리오균의 유전형인 임상형(rrnB/vcgC)과 환경형(rrnA/vcgE)을 확인하였다.
MLST
패혈증비브리오균의 MLST는 10개의 house-keeping 유전자 - glp (glucose-6-phosphate isomerase), gyrB (DNA gyrase, subunit B), mdh (malate-lactate dehydrogenase), metG (methionyl-tRNA synthetase), purM (phosphoribosylaminoimidazole synthetase), dtdS (threonine dehydrogenase), lysA (diaminopimelate decarboxylase), pntA (transhydrogenase alpha subunit), pyrC (dihydroorotase), tnaA (tryptophanse)-를 이용하여 sequence type (ST)을 분류하였다 (26, 27). PCR에 사용된 primer와 반응조건은 PubMLST (https://pubmlst.org/organisms/vibrio-vulnificus)에 제시된 방법을 사용하였다 (28). 전기영동으로 PCR 산물의 적합한 증폭을 확인한 후, 증폭산물의 염기서열은 ㈜마크로젠에 의뢰하여 분석하였다. 염기서열 정보는 SeqMan 프로그램(DNASTAR Inc, USA)으로 DNA assembly를 수행하여 편집한 후, 최종 염기서열을 확인하였다. 패혈증비브리오균 PubMLST 데이터베이스를 통해 ST를 확인하였다.
계통분석
균주들 간의 상동성을 확인하기 위해 BioEdit 프로그램을 이용하여 MLST 유전자 10개 각각에 대한 염기서열을 분석하였다. 패혈증비브리오균 77주의 MLST 결과를 glp, gyrB, mdh, metG, purM, dtdS, lysA, pntA, pyrC, tnaA 유전자 순으로 이은 4,326 bp 크기의 염기서열을 병렬 정렬한 후 계통분석에 사용하였다. 계통도는 MEGA 11 프로그램의 Neighbor-Joining 방법을 이용하여 작성하였으며, 계통수의 신뢰도 확보를 위해 1,000회의 부트스트랩(bootstrap)을 시행 후 신뢰 수준 50% 이상만 나타내었다.
RESULTS AND DISCUSSION
임상 특성
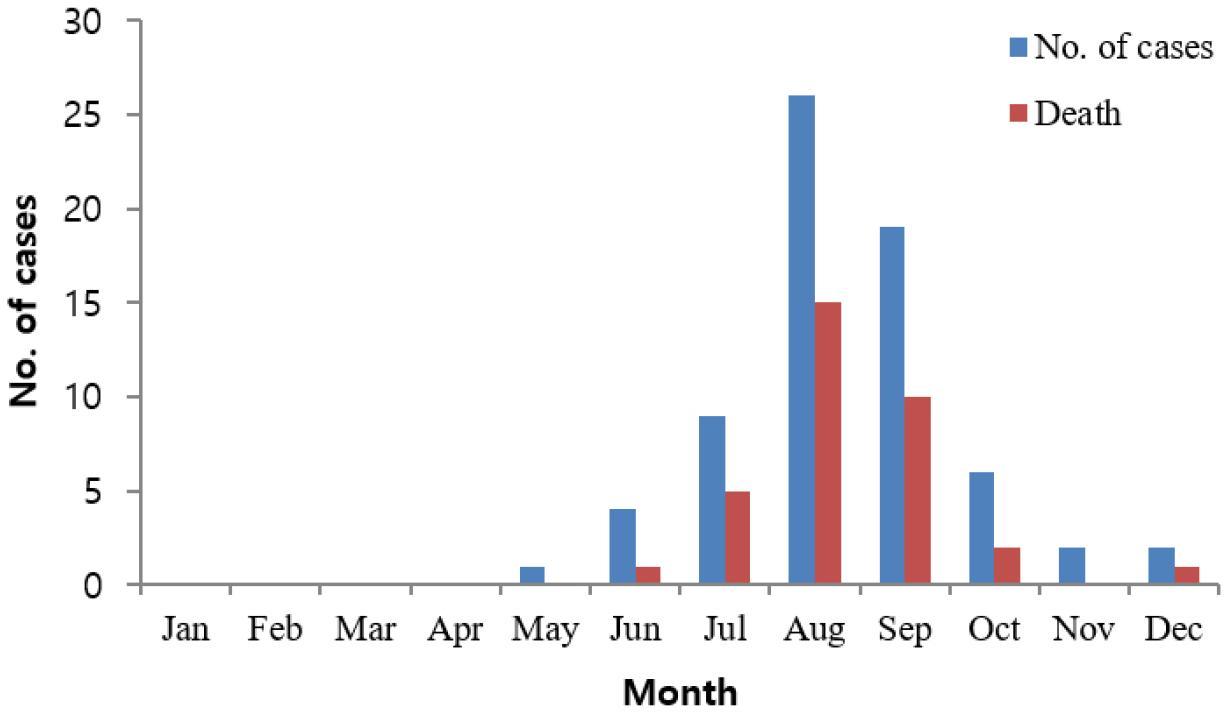
2014년부터 2022년까지 전라남도에서는 총 69건의 비브리오패혈증이 보고되었으며, 이 기간 동안 인구 10만 명당 평균 발생률은 0.41±0.21명으로 전국에서 가장 높은 발생률을 보였다 (7). 9년간 전라남도 평균 환자 발생건수는 7.7명이며, 사망자는 34명으로 49.3%의 치사율을 나타내었다 (Fig. 1). 비브리오패혈증의 유병률 증가는 온도와 상관관계가 있기 때문에(29, 30), 환자 발생과 계절성은 높은 상관성이 있다. 월별 발생은 8월에 가장 많았고(n=26, 37.7%), 그 뒤를 이어 9월(n=19, 27.5%), 7월(n=9, 13.0%) 순이었으며, 1~4월에는 전라남도에서 보고된 사례는 없었다. 연구에 사용한 균 분리 환자의 평균 연령은 64.7세(43-87)로, 남성의 비율은 77.1%(27명)이었으며, 전라남도 전체 환자군은 평균 연령이 63.9세(13-96), 남성 비율 76.8%(53명)로 비슷한 인적 특성을 보였다.
환경 특성
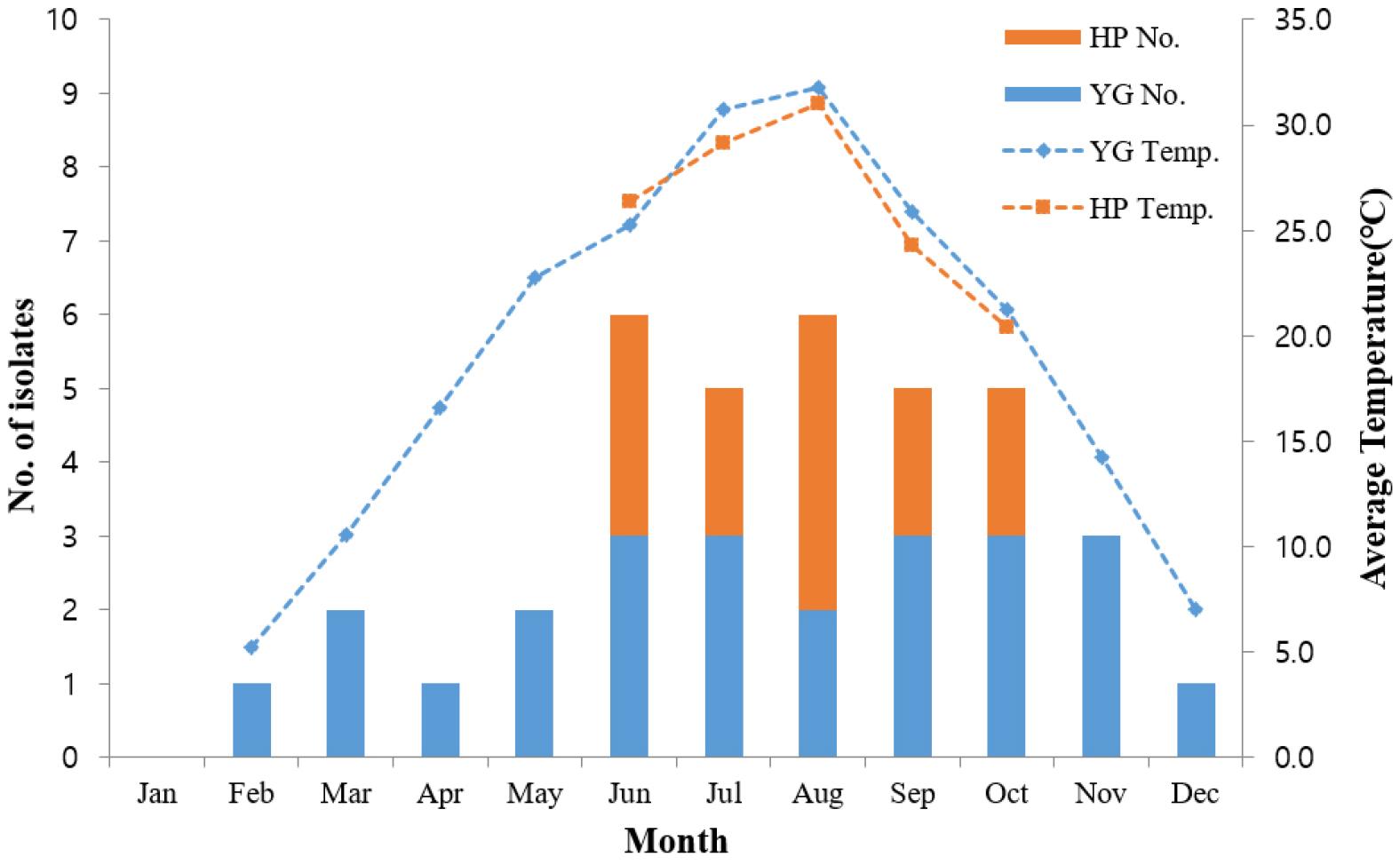
2022년 연중 비브리오 감시를 수행한 결과, 전라남도 영광과 함평의 해수와 갯벌에서 각각 23건, 14건의 패혈증비브리오균을 분리하였다. 검체 채취 시 측정한 온도는 해양환경에서의 패혈증비브리오균의 분리와 상관관계가 가장 높은 환경인자로 (31), 평균 검체 온도는 24.6℃였다. 영광지역에서 분리한 24건은 1월을 제외하고 연중 분리되었으며, 평균 검체 온도는 23.1℃였다. 함평지역에서 분리한 13건의 평균 검체 온도는 27.0℃로, 따뜻한 6월에서 10월 사이에 분리되었기 때문에 영광지역보다 평균 검체 온도가 더 높았다 (Fig. 2). 환자가 발생하기 시작하는 5월부터 해양환경의 온도는 20℃ 이상으로 상승하기 시작하여, 영광지역은 7월(33.3℃), 함평지역은 8월(32.3℃)에 가장 높은 온도를 보였다. 두 지역 모두10월부터 온도가 20℃ 이하로 감소하기 시작했으며, 가장 낮은 온도는 영광지역은 2월(5.2℃), 함평지역은 10월(18.9℃)에 관찰되었다. MLST 분석을 통해 영광지역 저온(5.2~16.6℃) 환경에서 분리한 5주가 동일한 ST를 보여, 2020~2021년 영광지역 저온(3.7~8.0℃)의 해수에서 분리한 5주를 추가로 연구에 포함시켰다.
임상 및 환경분리균주의 유전형 확인
16S rRNA유전자와 독성상관유전자를 이용한 유전형 분석에서 총 77주 중 62주(80.5%)는 임상형(rrnB/vcgC)이었다. 분리 출처와 관계없이 임상형의 비율이 임상분리균주에서는 80.0%(28주), 환경분리균주에서는 81.0%(34주)로 비슷하게 나타났다 (Table 3). 이전 연구에서도 굴에서 분리한 균주보다 해수에서 분리한 균주에서 임상형의 비율이 높았으며 (32), 2018년 국내 병원성 비브리오균 조사에서도 해수에서 분리한 패혈증비브리오균의 임상형 비율이 89.6%로 높은 수준을 보였다 (33). 환경형(rrnA/vcgE)은 모두 6주에서 확인되었으며, 환경분리균주에서 4주, 임상분리균주에서 2주이었다. 유전형을 나누는 두 유전자가 불일치한 경우는 모두 5건으로, rrnA/vcgC 유형이 4주, rrnB/vcgE 유형이 1주 있었다. 16S rRNA 유전자가 증폭되지 않은 4주는 독성상관유전자 유전형 검사에서 모두 vcgC로 확인되었다.
Table 3.
Genotypes of V. vulnificus isolates
| Samples | Genotype of vcg | No. of isolates | |||
|---|---|---|---|---|---|
| rrnA | rrnB | UCa | Total | ||
| Clinical | vcgC | 4 | 28 | 1 | 33 |
| vcgE | 2 | - | - | 2 | |
| Environmental | vcgC | - | 34 | 3 | 37 |
| vcgE | 4 | 1 | - | 5 | |
임상 및 환경분리균주의 ST 분석
연구에 사용된 77주의 패혈증비브리오균은 총 59개의 ST로 분류되었다(Table 4). 59개의 ST중 50개(84.7%)는 하나의 균주에서만 나타났으며, 두 균주 이상이 동일한 ST를 가지는 경우는 9개로 유전학적 다양성이 매우 높았다. 본 연구를 통해 기존 보고된 allele 및 ST와는 다른 30개의 allele 서열-glp(4), gyrB(2), mdh(2), metG(4), purM(3), dtdS(7), lysA(5), pntA(1), pyrC(1), tnaA(1)-과 37개의 ST를 신규로 발견하였다 (Table 4). 분리 출처에 따라 살펴보면, 임상분리균주 35주는 32개의 ST으로 나뉘었고, 그 중 16개(50%)가 새로운 ST이었다. 환경분리균주 42주는 29개의 ST으로 나뉘었고, 그 중 22개(75.9%)가 새로운 ST이었다. 두 균주 이상이 가지는 9개의 ST를 살펴보면, 임상분리균주에서만 나타나는 ST는 ST294와 ST655이었다. 환경분리균주에서만 나타나는 ST는 ST397, ST631, ST637, ST646으로 패혈증비브리오균 분리가 활발한 시기(5월~10월)에 같은 지역에서 중복되어 나타났다. 영광지역에서 분리한 환경분리균주 5주가 공유한 ST629는 2~4월, 그리고 12월에 저온(5.2-16.6℃)의 환경분리균주라는 공통점이 있었다. 추가로 2020~2021년 저온(6.2~8.0℃)의 환경에서 분리한 해수분리 5주를 분석한 결과, ST629로 총 10주의 환경분리균주가 동일한 ST를 나타냈다. 생쥐 모델을 통해 임상과 굴에서 분리한 패혈증비브리오균의 독성 변수들을 확인한 최근 연구 결과에서는 추운 계절(10~4월)에 분리한 균주가 따뜻한 계절(5~9월)에 분리한 균주보다 더 높은 간 감염을 일으키고, 중증도의 대용치(as a proxy for illness severity)인 쥐의 온도가 낮아짐을 유의미하게 확인하였다 (16). 임상과 환경분리균주들 간의 MLST 분석 결과 매우 다양한 ST가 나타나는 가운데, 저온의 환경에서 분리한 10주가 공통적으로 공유한 ST629 균주에 대한 독성 연구는 추가적으로 필요할 것으로 생각된다.
Table 4.
Sequence types of 77 V. vulnificus isolates
패혈증비브리오균의 계통 분석
임상검체와 환경에서 분리한 패혈증비브리오균 77주의 MLST 서열(4,326 bp)을 이용한 계통 분석 결과, 이전 연구들과 마찬가지로 두 개의 계통으로 나눌 수 있었다 (34, 35, 36). 두 계통은 분리 출처에 따른 분류가 아닌 유전형에 따라서 계통학적으로 나뉘어졌다 (Fig. 3). 유전형에 따라 나뉜 계통도를 임상형은 계통 C, 환경형은 계통 E로 본 연구에서 임의적으로 분류하였다. 계통 C에는 임상형(rrnB/vcgC) 균주와 JNIHE_C23을 제외한 비특정 유전형 8주가 모두 포함되었으며, 대표적인 참조균주인 CMCP6 (ST17), MO6-24/O (ST1), YJ106 (ST22)을 모두 포함하였다. 계통 E는 환경형(rrnA/vcgE) 균주들에서만 관찰되었으며, 임상분리균주 2주와 환경분리균주 4주가 포함되었다. 임상분리균주 JNIHE_C23은 유전형 rrnA/vcgC로 비특정 유전형을 가진 9주 중 유일하게 두 계통에 속하지 않았다.
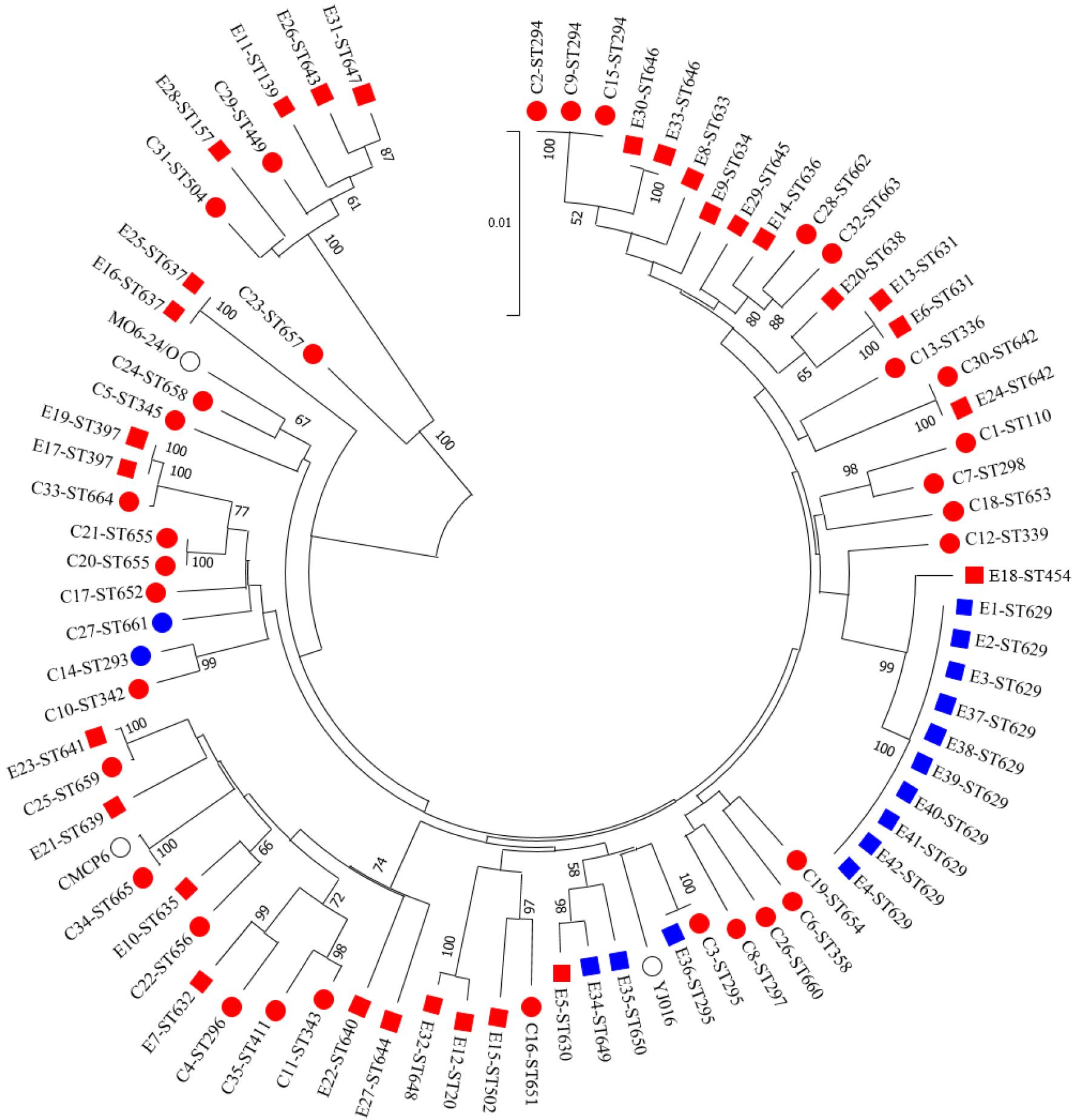
Fig. 3
Phylogenetic neighbor-joining circular tree of 77 V. vulnificus isolates and three reference strains by the concatenated sequences of 10 house-keeping genes used in MLST analysis. Circle (○) and square (□) represent clinical and environmental isolates, respectively. Blue and red color represent the isolates from the cold season (November to April) and the warm season (May to October), respectively. No color was shown in three reference strains.
계통 분석과 균주 특성의 연관성 분석
77주의 패혈증비브리오균 중 70주(90.9%)가 계통 C로 분류되어 임상검체와 환경분리균의 계통 차이는 없었다. 따라서 계통 C에 속한 환경분리균주가 계통 E의 환경분리균주 보다 독성이 높다고는 할 수 없다. 환경분리균주 관련 정보인 분리 지역 및 검체 종류에 따른 연관성은 계통도 내 산발적 분포로 인해 발견하기 어려웠다. 임상과 환경분리균주들 간의 계절적 시기를 환경 검체 채취 시 온도 20℃를 기준으로 따뜻한 계절(5~10월)과 추운 계절(11~4월)로 나누었다. 따뜻한 계절에 분리한 균주는 전체 균주의 80.5%(62건)로 임상분리 33주와 환경분리 29주가 포함되었으며, 이들 균주는 두 계통에 산발적으로 분포하였다. 추운 계절에 분리한 임상분리 2주와 환경분리 13주는 모두 계통 C에 속하였다 (Fig. 3). 패혈증비브리오균의 발생 및 분리는 계절성이 뚜렷하기 때문에, 추운 계절(11~4월)에 임상과 환경에서 분리된 균에 대한 연구는 아직 부족하다. 본 연구의 임상 및 환경분리균주 관련 특성과 계통 분석 결과, 균 분리 온도와 패혈증비브리오균의 독성에 대한 추가적인 연구가 필요할 것으로 생각된다. 또한 지구온난화에 따라 병원성 비브리오균에 대한 위험도가 높아진 가운데, 비브리오패혈증 발생이 없던 지역에서도 질병이 발생하고 있다 (37, 38). 다양한 환경에서 분리되는 패혈증비브리오균이 동일한 병원성을 가지고 있지 않으므로 독성이 높은 균주를 신속하고 정확하게 구별할 수 있는 분자 역학적 표지자가 필요하다.
CONCLUSION
비브리오패혈증의 발생률이 전국에서 가장 높은 전라남도에서 분리한 77주의 임상 및 환경분리균주를 대상으로 유전형과ST를 분석하였으며, MLST 분석에 이용한 유전자의 염기서열을 바탕으로 균주의 계통 분석과 균 분리 계절 및 기온의 연관성을 조사하였다. 연구에 사용한 77주는 MLST법으로 모두 59개의 ST로 분류되었으며, 50개(84.7%)의 ST는 하나의 균주에서만 확인되어 다양한 클론의 패혈증비브리오균이 분포하고 있었다. 3년간(2020-2022년) 영광지역의 저온(5.2-16.6℃) 환경에서 분리한 10주는 모두 ST629로 동일하였다. 유전형의 분포는 임상형(rrnB/vcgC)의 비율이 임상분리주와 환경분리주에서 비슷하였으며, 계통학적 위치는 유전형에 따라 임상형(계통 C)과 환경형(계통 E)으로 나눌 수 있었다. 추운 계절(11~4월)에 분리된 15주의 임상 및 환경분리균주는 모두 계통C에 속하였다. 이러한 연구 결과는 우리나라에서 분리되는 패혈증비브리오균의 계통학적 분석의 기초자료로 활용될 수 있으며, 비브리오패혈증의 감염 관리 및 예방 대책 수립의 기초자료로도 활용될 수 있을 것이다.
