INTRODUCTION
The genus Mycobacterium includes the Mycobacterium tuberculosis (Mtb) complex, Mycobacterium leprae, and non-tuberculous mycobacteria (NTM) (1). Tuberculosis (TB) remains a leading cause of mortality worldwide, necessitating ongoing research into vaccines and pathogenic mechanisms to enhance control measures and ultimately eradicate TB (2, 3). Despite the significant focus on TB, there is growing concern regarding the rising incidence of NTM infections (4, 5, 6). Unlike Mtb, which survives in the human body and is transmitted between people, NTM are widely distributed in nature, including rivers and soil, and are not usually transmitted between people through respiratory infections (7). Although TB incidence is declining in industrialized countries, NTM infections are steadily increasing (5). However, because NTM include many more species than Mtb, research on each of these bacteria has been relatively limited compared to TB, despite the potential risks and applications associated with NTM (8). Therefore, further study is needed on the pathogenicity, treatment methods, and potential applications of each NTM strain.
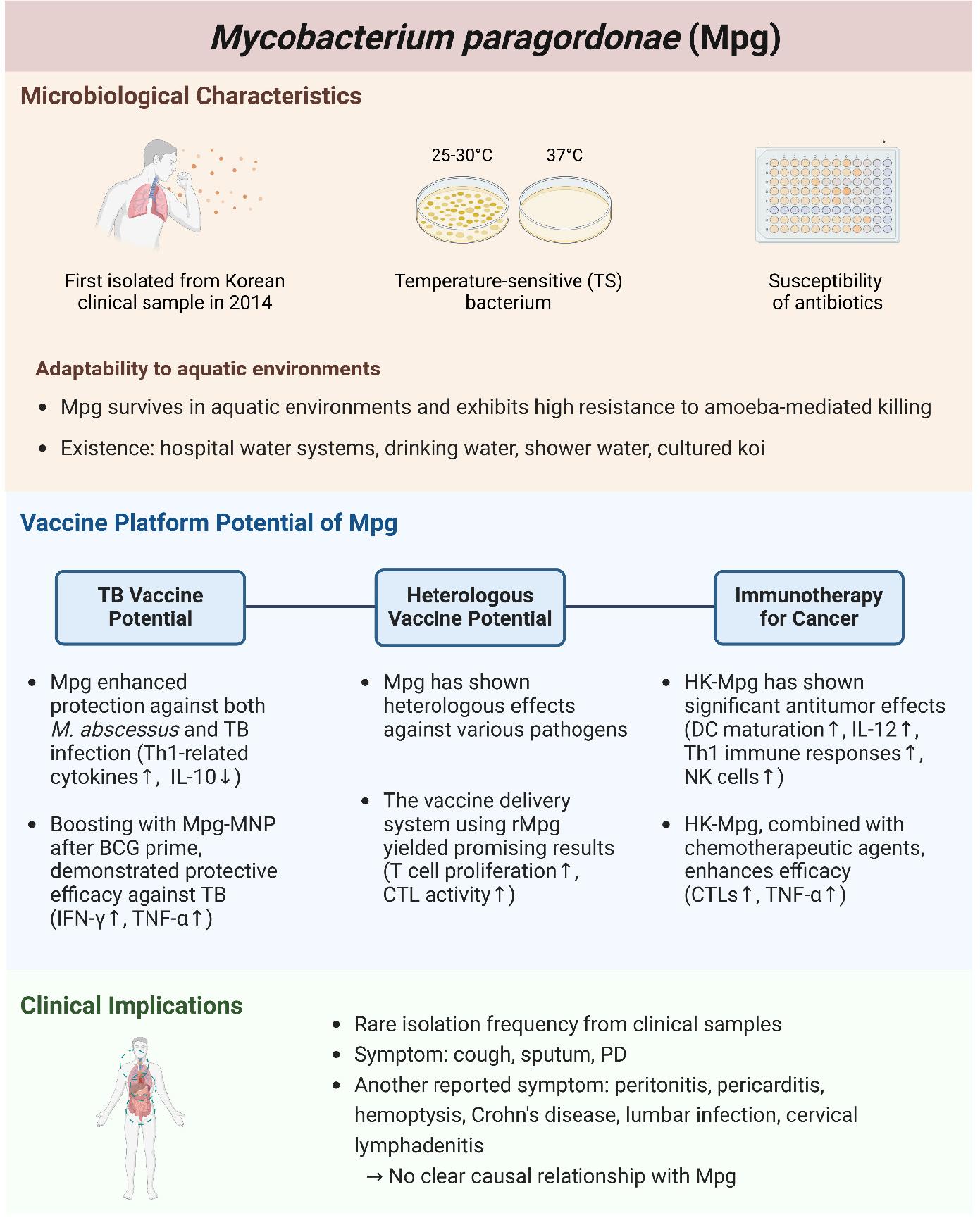
Mycobacterium paragordonae (Mpg) is a recently identified species first described in 2014 (9). Mpg presents notable potential as a vaccine candidate due to its unique characteristics, including temperature sensitivity and adaptability to diverse environments, including clinical settings (10, 11, 12, 13, 14). It has been proposed as a novel live vaccine to prevent mycobacterial infections and has demonstrated promising results in inducing immune responses in various studies (10, 14). Furthermore, one of the characteristics of BCG heterologous immunity—which acts as innate immune memory—has also been observed in Mpg, activating the innate immune response by leveraging type I interferons (IFN-I) from dendritic cells (DC), suggesting its potential application across various disease fields (13, 15). Mpg, which demonstrates these beneficial properties, exhibits a more adaptive intracellular lifestyle within amoeba, which are aquatic protozoa, compared to other mycobacteria, leading to its reporting in water-related infections (16). Mpg rarely appears in clinical case studies and a few infection cases will be discussed in this review (17, 18, 19, 20, 21).
This review aims to provide a comprehensive summary of the current research status of Mpg. By compiling published literature, we aim to highlight key findings and emphasize the potential applications of Mpg in various research fields, including clinical applications and vaccine development.
PHENOTYPIC AND PHYLOGENETIC CHARACTERISTICS OF MPG
Mpg was first isolated from a clinical sample of a patient’s sputum with symptomatic pulmonary infections during an investigation involving the hsp65-sequence-based identification of Korean clinical isolates. The strain, designated as 49061T, was initially suspected to be M. gordonae due to its phenotypic similarities. Although strain 49061T shared several phenotypic traits with M. gordonae, including being a rod-shaped and acid-fast bacterium with curved bacilli without spores or filaments observable by microscopy, there were notable differences. Unlike M. gordonae, which grows optimally at 37°C and produces yellow or white colonies, strain 49061T exhibited optimal growth at lower temperatures of 25-30°C and failed to grow at 37°C on commonly used Middlebrook 7H10 agar slants. The colonies of strain 49061T were smooth and orange in color, distinguishing it from M. gordonae(9).
In some studies, it has been reported that Mpg grows at 37°C on 7H11 agar, Lowenstein-Jensen (LJ) media, and in the BD BACTEC MGIT 960 system (22, 23, 24). Considering this, it is likely that these differences may be attributable to the composition of each medium. First, 7H11 agar is an enriched medium that includes casein hydrolysate added to 7H10 agar, providing additional nutrients (25). LJ media primarily contains eggs, which supply proteins and lipids, while the BD BACTEC MGIT 960 system utilizes modified Middlebrook 7H9 liquid media with casein peptone (26, 27). Overall, the studies reporting Mpg growth at 37°C appear to involve media with higher nutrient content, such as casein, compared to 7H9 broth or 7H10 agar, where Mpg has previously shown temperature sensitivity. However, the specific nutrient components that influence the temperature-sensitive (TS) growth characteristics of Mpg have yet to be determined. Further research on this issue could advance our understanding not only of Mpg but also of TS bacteria more broadly.
Genomic analysis was performed to definitively determine its taxonomic status. Phylogenetic analyses based on 16S rRNA, hsp65, and rpoB gene sequences indicated it as a distinct species. DNA-DNA hybridization assay also supported this classification, leading to the recognition of strain 49061T as a novel species, formally named Mycobacterium paragordonae (type strain 49061T = JCM 18565T = KCTC 29126T) in 2014 (9).
Biochemical analysis revealed unique profiles for strain 49061T, demonstrating tolerance to thiophene-2-carboxylic acid hydrazide (TCH) and lack of growth on media containing high concentrations of p-nitrobenzoate (PNB), 5% sodium chloride, picric acid, or ethambutol. Additionally, strain 49061T didn’t accumulate niacin or reduce nitrate, which are typical reactions for M. gordonae (9).
ANTIBIOTIC SUSCEPTIBILITY OF MPG AND ITS USE FOR DRUG SCREENING
Mpg shows high susceptibility to a broad spectrum of antibiotics, making it a valuable organism for antibiotic screening. The high susceptibility of Mpg allows it to efficiently eliminate ineffective compounds, making it particularly useful in the early stages of drug screening. This advantage is also important when only small amounts of compounds are available for testing.
In studies confirming the susceptibility of Mpg to various antibiotics using the SLOMYCO Sensititre panel, it showed significantly lower minimum inhibitory concentrations (MICs) for antibiotics such as ciprofloxacin, clarithromycin, isoniazid, moxifloxacin, rifampin, and streptomycin compared to closely related species like M. marinum and M. gordonae. The MICs for Mpg were being 4-8 times lower, highlighting its potential as a model organism for antibiotic research (28).
Utilizing its high sensitivity to antibiotics, a recombinant Mpg strain expressing a luciferase reporter gene (rMpg-LuxG13) was developed to improve the efficiency and accuracy of antibiotic screening assays. The rMpg-luxG13 facilitates real-time monitoring of bacterial viability through luminescence measurements, enabling high-throughput screening. This drug screening model was developed using both amoeba-based and in vitro direct application systems. The amoeba-based system was conducted by infecting Acanthamoeba castellanii with the rMpg-LuxG13 strain to assess their interactions. The amoebae infected for 2 hours at 28 ˚C were cultured in PYG medium containing 10 μM amikacin to remove extracellular bacteria, and subsequently used for drug screening. Studies have shown that antibiotics such as rifampin, isoniazid, clarithromycin, and ciprofloxacin effectively inhibit the growth of rMpg-LuxG13, demonstrating the strain’s utility in antibiotic screening and drug discovery (28).
POTENTIAL OF MPG AS A LIVE TB VACCINE
The unique characteristics of Mpg, such as its susceptibility to various antibiotics and TS growth, highlight its potential as a safe live vaccine candidate. Unlike BCG, Mpg retains the RD1 region, which is crucial for presenting antigens related to immune system resistance and evasion (16). When BCG was complemented with key RD1 genes, the complemented BCG exhibited enhanced protection against Mtb infections in preclinical models (29). Furthermore, Mpg harbors all five ESX loci (ESX-1, ESX-2, ESX-3, ESX-4, and ESX-5) found in Mtb, which may contribute to enhanced protection against TB (16).
Studies have demonstrated that Mpg can elicit robust immune responses surpassing those of BCG in mouse models. Mpg activates Th1-related cytokines such as IFN-γ and IL-12, essential for an effective immune response against mycobacterial infections like M. abscessus and Mtb, and reduces IL-10 expression associated with an anti-inflammatory response compared to BCG. Additionally, Mpg enhances DC maturation, evidenced by elevated levels of MHC II, CD40, CD80, and CD86, increasing their migratory capacity due to high CCR7 expression and ability to proliferate T cells, suggesting significant implications for developing more effective TB vaccines (10).
Mpg also shows promise as a booster vaccine. In countries where TB is prevalent, most individuals have completed BCG vaccination for TB prevention (30). Therefore, there is considerable interest in booster vaccines that can enhance the weakened immune response in adults following the initial BCG prime. When used as a booster following a BCG prime, Mpg elicited higher cell-mediated immune responses, such as elevated IFN-γ expression and reduced lung inflammation in mouse models (10). Additionally, the use of Mpg in biodegradable microneedle patches (MNP) has further demonstrated its potential as a non-invasive vaccine. Mpg-loaded MNPs applied to mouse skin induced significant immune cell recruitment into the dermis, which was not seen with subcutaneous administration (14). Among the recruited cells, there was a substantial increase in MHC II+ langerin+ cells and Ki-67+ langerin+ cells, which possess migratory capacity and play a key role in initiating T cell-mediated immune responses (31, 32). As a result, higher proportions of IFN-γ and TNF-α-secreting cells were observed in lymph nodes, highlighting its efficacy as a vaccine delivery method. Following a BCG prime, boosting with the Mpg-MNP method demonstrated protective efficacy against the virulent Mtb K-strain, a member of the Beijing strain family (14, 33). These results suggest that Mpg has potential as a novel live attenuated bacterial TB vaccine that could serve as an improved prime vaccine replacing BCG and as a booster vaccine.
HETEROLOGOUS VACCINE POTENTIAL AGAINST DIVERSE PATHOGENS AND USE OF RECOMBINANT MPG FOR VACCINE DELIVERY SYSTEMS
Heterologous vaccination strategies, which enhance innate immune responses to provide broad protection against various pathogens, have shown promise with Mpg (15). Like BCG, which is well-known for its heterologous vaccination effect, Mpg has demonstrated the ability to elicit strong immune responses not only against TB but also against other pathogens like Candida albicans, Staphylococcus aureus, and Escherichia coli (34, 35, 36, 37, 38, 39, 40). This broad-spectrum protection underscores Mpg’s potential as a versatile vaccine candidate. Notably, in SCID (Severe Combined Immunodeficient) mice, which are B-and T-cell deficient, Mpg demonstrated higher protective efficacy compared to BCG, confirming its potential as a stronger heterologous vaccine. Its heterologous vaccine efficacy is mainly attributed to the presence of cyclic di-GMP found in live Mpg, which can play a crucial role in the activation of innate immune cells via induction of IFN-I production (13). The cyclic di-GMP acts as a viability-associated pathogen-associated molecular pattern (vita-PAMP) to induce IFN-I through a stimulator of interferon genes (STING)-mediated endoplasmic reticulum stress response in DC (41). As a result of IFN-I production by DCs, the adaptive memory properties of NK cells are activated, leading to effective immune responses even to secondary stimuli (13, 42, 43).
Given its ability to induce strong immune responses, Mpg has been used to create recombinant forms that express heterologous antigens. Research into vaccine delivery systems using these recombinant Mpg strains has yielded promising results. For instance, recombinant Mpg strains expressing HIV-1 p24 antigens have shown higher T cell proliferation and cytotoxic T lymphocyte (CTL) activity compared to recombinant BCG expressing the same antigens (12). Similarly, immunization with recombinant Mpg expressing the receptor-binding domain (RBD) of SARS-CoV-2 has shown potential in activating immune responses against COVID-19, reducing viral load in SARS-CoV-2-infected cell models, and enhancing in vitro DC activation, indicating its utility in developing vaccines for various infectious diseases (11).
IMMUNOTHERAPY FOR CANCER
Mpg has shown significant promise in cancer immunotherapy as well as in treating infectious diseases. Heat-killed Mpg (HK-Mpg) has demonstrated notable antitumor effects in preclinical models, reducing tumor incidence and size and improving survival rates. These effects are attributed to its ability to strongly induce immune responses by promoting DC maturation and IL-12 production, which are critical for skewing the immune response towards a Th1 profile and activating natural killer (NK) cells (44).
Combining HK-Mpg with conventional chemotherapeutic agents such as cisplatin has shown enhanced efficacy, suggesting that Mpg can potentiate the effects of existing cancer treatments. HK-Mpg has been particularly effective in inducing T-cell responses, including the activation of CTLs and the production of TNF-α, both of which are pivotal in targeting and eliminating cancer cells (44). The ability of HK-Mpg to induce a strong immune response, combined with its synergistic effects when used alongside chemotherapy, positions it as a promising candidate for cancer immunotherapy and an adjunctive immunotherapy approach.
ENVIRONMENTAL ADAPTABILITY AND CLINICAL IMPLICATIONS OF MPG
Mpg has demonstrated remarkable environmental adaptability, particularly in aquatic environments. This adaptability is primarily attributed to its unique genomic features and survival mechanisms that enable it to thrive under diverse conditions. Below, we explore various documented cases of Mpg presence in water systems, providing a detailed overview of its survival strategies and public health implications.
Genomic Characteristics of Mpg
Mpg can survive in a variety of aquatic environments, which has been proven in several studies (24, 45). The increasing frequency of isolation of Mpg from aquatic systems has led to the hypothesis that free-living amoeba systems are likely natural reservoirs of Mpg. A study comparing the intracellular viability of Mpg, M. gordonae, and M. marinum within A. castellanii by counting colony-forming units (CFUs) over seven days showed that Mpg survived at significantly higher levels than M. marinum and M. gordonae, suggesting that Mpg may possess enhanced resistance to phagocytic process after infecting amoeba (16).
A significant feature of the Mpg genome is its massive acquisition of Type VII Secretion System (T7SS)-related genes, which is predicted to have influenced its adaptation to an intracellular lifestyle within environmental predators such as free-living amoebae (16). The genome of Mpg contains all five types of ESX loci, similar to pathogenic species like TB. However, the ESX-2 locus of Mpg is split into two separate regions within its genome, unlike the contiguous arrangement in TB (46). Additionally, Mpg possesses plasmid-encoded ESX systems, including ESX-P5 on plasmid pMpg-1 and an ESX-2-like system on plasmid pMpg-2. This unique genomic configuration, resulting from lateral gene transfer and genomic rearrangements, highlights the adaptation of Mpg to free-living amoebae and its enhanced intracellular survival capacity (16).
Documented Cases in Water Systems
Several studies have demonstrated that Mpg is found in hospital water systems, drinking water, shower water, and even cultured koi (Cyprinus carpio) (18, 19, 20, 24, 47, 48, 49). The consistent detection of Mpg in hospital water sources suggests that it can thrive in aquatic environments, adapting to various conditions and demonstrating high survival rates in such water systems. These findings underscore the importance of Mpg in infection control and water quality management.
Water Supply System: The increasing frequency of clinical isolation of NTM after hospital reconstruction and renovation at Miyazaki University Hospital in Japan prompted an analysis of the potential relationship between NTM and the hospital water supply system. During the outbreak investigation from April to July 2015, Mpg was found in 26.1% of 69 environmental samples. The investigation revealed a significant presence of NTM in recently replaced aerators/rectifiers. Removing these devices and preventing patients from consuming tap water resulted in a decrease in Mpg-positive samples (19).
Shower Water and Indoor Air: A study conducted in the United States showed that shower water contributes significantly to the bacterial community in indoor air and transmits NTM during showering. The dominant NTM species in shower water were M. mucogenicum and Mpg. Mpg accounted for 74.5% and 64.5% of NTM species during showers using rain showerheads and massage showerheads, respectively. The aerosolization of NTM during showering poses a potential infection risk. To minimize exposure, it is recommended to clean showerheads regularly and use water filters (47).
Hospital Water Sources in Iran: A review paper summarizing NTM finding in hospital water sources from various cities in Iran from 2016 to 2020 reported a varied presence of NTMs, including Mpg. Mpg was recorded in three studies, with a prevalence of 21.111% in samples from Isfahan, 6.251% in samples from Tabriz, and 2.321% in samples from Tehran (18, 24, 48). The high prevalence of NTM, including Mpg, highlights the need for rigorous water quality monitoring (20).
Cultured Koi Carp: Mpg has been reported to be isolated from naturally occurring carp exhibiting lethargy, weakness, and pinhead-shaped bodies in aquaculture ponds in Niigata, Japan. Mpg, along with two other Mycobacterium species, caused mycobacteriosis in carp. This suggests that Mpg may be transferred from aquatic environments to aquaculture environments, underscoring the need for regular monitoring and the development of effective antibiotics and vaccines (49).
Clinical Implications
Mpg infection in humans is known to be relatively rare. According to a cohort study in China, Mpg was isolated from only 8 out of 1,730 NTM samples (0.46%) (23). In most cases, it presents with symptoms similar to common NTM infections, such as coughing, sputum production, and pulmonary disease. However, several cases have been reported with different symptoms (21, 50, 51, 52, 53, 54, 55). Documented clinical cases include peritonitis, pericarditis, hemoptysis, Crohn’s disease, lumbar infection, and cervical lymphadenitis (Table 1). These occurrences are sporadic, and most reported cases involve immunocompromised individuals or specific hospital settings. The following are reported Mpg clinical infection case and their symptoms.
Table 1.
Reported Mpg Clinical Infection Cases
|
Gender /Age | Pre-existing Conditions | Symptoms | Diagnostic Methods | Treatment Regimen | Treatment Duration | Outcome | Country/ Publication year | Ref |
|---|---|---|---|---|---|---|---|---|
|
Male /55 | End-stage kidney disease due to diabetic nephropathy | Abdominal pain, cloudy peritoneal dialysate effluent | PD effluent culture, Mycobacteria growth indicator tube | Amikacin, Azithromycin, Ethambutol | 10 weeks | Died of ischemic heart disease |
Hong Kong /2017 | (21) |
|
Female /60 | Hypothyroidism, dyslipidemia | Chest pain, hypoxia | Echocardiogram, CT scan, Pericardiocentesis, Mycobacterial culture | Ethambutol, Ciprofloxacin, Doxycycline | 12 months | Stable |
Canada /2023 | (50) |
|
Male /44 | Crohn’s Disease | Abdominal pain, non-bloody diarrhea | Colonoscopy, CT scan | Mesalamine, Isoniazid, Rifampin | 5 months | Improved |
United States /2022 | (51) |
|
Female /54 | None | Malaise, hemoptysis, diffuse rhonchi in lungs | Chest X-ray, CT, Bronchoalveolar lavage, Acid-fast bacilli culture | Azithromycin, Ceftriaxone | No Data | Lost follow-up |
United States /2019 | (52) |
|
Male /53 | None | Lumbar pain | MRI, Biopsy, Culture |
Rifampin, Ethambutol, Azithromycin, Moxifloxacin | 1 month | Improved |
China /2021 | (53) |
|
Female /20s | None | Enlarged, mobile right cervical lymph node, pain on swallowing, slightly tender | Biopsy, bloodwork, ultrasound, CT scan, culture | Amoxicillin/clavulanate, ciprofloxacin, ertapenem, clindamycin, clarithromycin | 3 months | Resolved |
Canada /2022 | (54, 55) |
Peritonitis: A 55-year-old man on continuous ambulatory peritoneal dialysis (CAPD) developed peritonitis caused by Mpg. He presented with cloudy peritoneal dialysate and abdominal pain. Initial treatment with intraperitoneal cefazolin and cefepime was temporarily effective, but symptoms recurred. Mpg was identified after 2 weeks of incubation. Despite switching to intravenous antibiotics and removing the catheter, the patient did not improve significantly and ultimately succumbed to ischemic heart disease after 10 weeks (21).
Pericarditis: A 60-year-old woman with hypothyroidism and dyslipidemia presented with chest pain and hypoxia, diagnosed as Mpg-associated pericarditis with pericardial masses. Mpg was later found in the pericardial fluid. After 12 months of treatment with ethambutol, ciprofloxacin, and doxycycline, her symptoms and pericardial effusion significantly improved, highlighting the rarity and severity of Mpg infections in cardiac disease (50).
Crohn’s Disease: A 44-year-old man with right-sided abdominal pain and non-bloody diarrhea was diagnosed with Crohn’s disease (CD) after a colonoscopy. During treatment, a TB test was positive, and a chest X-ray showed fibronodular changes. Although the sputum acid-fast smear was negative, cultures identified Mpg. The initial RIPE regimen was used but switched to isoniazid and rifampin due to side effects. Significant improvement in colitis was observed following mycobacterial treatment, suggesting a potential link between Mpg and CD (51).
Hemoptysis: A 54-year-old woman presented with hemoptysis and bilateral lung abnormalities, initially treated for community-acquired pneumonia. Initial respiratory cultures and acid-fast bacilli tests were negative, but later cultures identified Mpg. Despite initial improvement with azithromycin and ceftriaxone, the patient was lost to follow-up, leaving the long- term outcome unknown. This case highlights the need to consider slow-growing mycobacteria in persistent hemoptysis (52).
Lumbar Infection: A case of lumbar infection in a patient was attributed to Mpg, manifesting as persistent back pain. The diagnosis was confirmed through imaging and cultures. The patient responded well to a combination of antimicrobial therapy, including ethambutol and moxifloxacin, resulting in significant recovery. This case underscores the ability of Mpg to affect the musculoskeletal system and the importance of appropriate mycobacterial treatment (53).
Cervical Lymphadenitis: A rare case of infectious cervical lymphadenitis, in which both Citrobacter freundii, MAC, and Mpg were positive on core biopsy, has been reported. The patient received multiple antibiotics concurrently and was managed conservatively without surgical intervention. Symptoms resolved after 3 days of antimycobacterial treatment (clarithromycin), followed up for 3 months (54, 55).
Mpg has been associated with several clinical cases of infection. However, in most instances, a definitive causal relationship has not been clearly established. Rapid and accurate identification of Mpg infections is crucial, as patients show significant improvement when treated with anti-tuberculosis drugs and antibiotics. When conducting NTM culture tests, it is essential to consider the possibility of slow-growing mycobacteria. Therefore, cultures should be maintained for a duration of more than a month to ensure accurate detection. Given the stable prognosis of Mpg infections, precise detection is vital for establishing effective management and prevention strategies.
CONCLUSION
Mpg exhibits unique characteristics and versatility, making it a promising candidate for vaccine development, immunotherapy, and antibiotic screening. Its robust immunogenicity, potential as a recombinant vaccine vector, and efficacy in heterologous vaccination strategies highlight its potential for developing new and improved vaccines and therapies against various diseases. The environmental adaptability and clinical implications of Mpg underscore the need for ongoing research and surveillance to develop effective management and prevention strategies (Fig. 1). Further research and clinical trials are essential to fully realize the potential of Mpg in addressing some of the most pressing challenges in medicine and public health.